



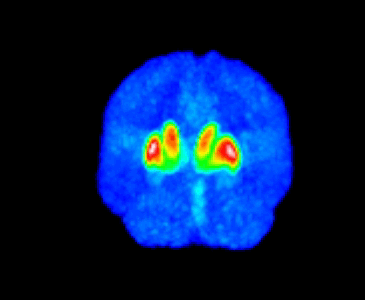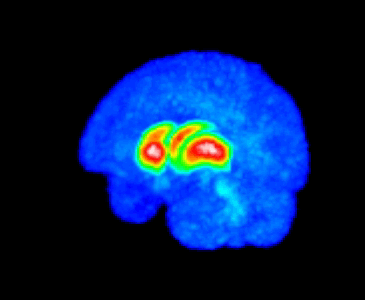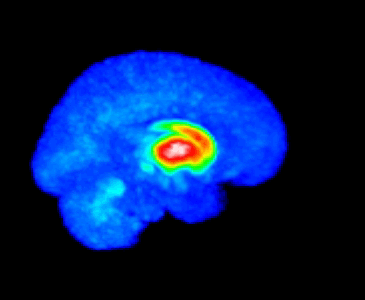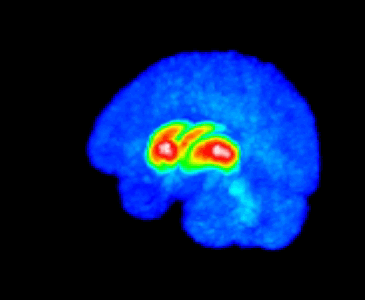
 The MMTI Lab uses human neuroimaging approaches such as Positron Emission Tomography (PET), functional Magnetic Resonance Imaging (fMRI), and Magnetic Resonance Spectroscopy (MRS), as well as pharmacological and other (e.g. repetitive transcranial magnetic stimulation; rTMS) interventions, to advance our understanding of the neural mechanisms of psychiatric, neurological, and substance use disorders, with a special emphasis on schizophrenia spectrum disorders and dopaminergic systems in the striatum.
The MMTI Lab uses human neuroimaging approaches such as Positron Emission Tomography (PET), functional Magnetic Resonance Imaging (fMRI), and Magnetic Resonance Spectroscopy (MRS), as well as pharmacological and other (e.g. repetitive transcranial magnetic stimulation; rTMS) interventions, to advance our understanding of the neural mechanisms of psychiatric, neurological, and substance use disorders, with a special emphasis on schizophrenia spectrum disorders and dopaminergic systems in the striatum.
Interested in Participating?
To learn about our studies and how you can participate please call 631.885.0814 or email MMTILab.SBU@gmail.com.
Multi-Modal Translational Imaging Lab (MMTI Lab)
Health Sciences Center (HSC)
101 Nicolls Road
Suite 087, Floor T10
Stony Brook, NY 11794
